Un nuovo indice potrebbe fornire una stima universale della diversità biologica
Conosciamo molto meglio alcuni pianeti del nostro sistema solare che gli ecosistemi marini della nostra Terra e le specie che li popolano. Ad esempio, solo da qualche anno la comunità scientifica ha acquisito maggiori informazioni biologiche sulle profondità oceaniche, ma molto resta ancora da scoprire. Sappiamo ancora poco della diversità biologica delle barriere coralline (come quella hawaiana della figura sotto), ma la stiamo perdendo a ritmi insostenibili a causa degli impatti umani su queste fragili comunità di organismi. Ciò che è, ormai, certamente chiaro è che la straordinaria biodiversità di questi ambienti marini, così come quella delle più esplorate foreste tropicali, ha estremo bisogno di essere monitorata, misurata e protetta.

Proprio per questo, l’identificazione di un indice universale di diversità biologica, che può essere applicato a ogni ricerca ed ecosistema, è uno degli obiettivi principali degli ecologi. La metrica più semplice, ovvero la ricchezza di specie, sebbene fornisca un numero esatto di specie in un campione, può essere poco istruttiva e altamente imprecisa, essendo soggetta ai cambiamenti casuali nella composizione delle specie più di qualsiasi altro indice. D’altronde, gli indici di diversità basati sull’abbondanza aggiungono l’informazione riguardante il numero di individui a quello della ricchezza delle specie, ma non sono in grado di rappresentare la vera “diversità” di una comunità.
Gli indici che incorporano una misura di eterogeneità (o distribuzione degli individui all’interno delle specie) a una di ricchezza sono chiamati “indici di diversità” e quelli di Shannon e Simpson (a volte chiamato Gini-Simpson) sono i più utilizzati. Tuttavia, questi non possono essere considerati indici di “vera diversità” perché entrambi sono altamente non lineari e i loro valori non sono comparabili in termini di numero effettivo di specie (e della loro relativa abbondanza) in una comunità.
Recentemente sono stati proposti due tipologie di indici, assoluti ed effettivi, che meglio stimano la biodiversità di una comunità o area. Gli indici di “diversità assoluta” sono basati sul rapporto statistico simile al procedimento di cattura, rilascio e ricattura (in inglese mark-release-recapture; MRR) per stimare la ricchezza aggiungendo un fattore di correzione al numero osservato di specie. Successivamente, Jost (2006) ha definito la “diversità effettiva” come il numero di specie ugualmente abbondanti, che equivalgono ai valori alpha 0, 1 e 2 dell’equazione di Hill (1973). Infatti, gli indici di ricchezza di specie, diversità di Shannon e diversità di Simpson sono casi speciali di misure di entropia. Questi indici sono stati “riassunti” da Renyi (1961) e poi “formalizzati” da Hill in una singola equazione che può aiutare a ottenere una descrizione precisa semplicemente cambiando un parametro alfa.
Gli indici di biodiversità assoluta
Uno degli indici di diversità assoluta più affidabili, che considera la rarità delle specie rappresentate da un individuo (chiamate singletons) e due individui (doubletons), è il Chao1, sviluppato da Anne Chao (Chao 1984). Chao2 è lo stesso indice applicato ai dati basati sull’incidenza. Chao1 stima il numero assoluto di specie in un campione, in base al numero di specie rare, come numero di specie realmente osservate sommato al rapporto tra singletons e doubletons. Il problema principale di questo stimatore è che è fortemente influenzato dalla dimensione del campione, quindi è importante garantire un campionamento adeguato per non sottovalutare la ricchezza. Inoltre, l’indice Chao1 fornisce una stima minima di ricchezza e assume che ci sia omogeneità tra i campioni. Sarebbe, quindi, inappropriato usarlo per confrontare la ricchezza stimata di siti che hanno una grande differenza nella loro composizione specifica.
Il numero effettivo di specie
Allo stesso tempo, i vantaggi di derivare il “numero effettivo di specie” dagli indici di diversità canonici di Shannon e Simpson hanno ottenuto recenti consensi. Questo perché la vera diversità di una comunità può anche essere considerata il numero di specie ugualmente comuni richieste per dare un determinato valore a un indice. Questo approccio ha un’interpretazione intuitiva. Infatti, dopo la conversione in unità comuni di diversità effettiva, è possibile eseguire un confronto dei valori calcolanti nella stessa area. Ciò rende possibile un trattamento unificato di tutti gli indici di diversità effettiva e di confrontare la vera diversità tra comunità supponendo che tutte le specie siano ugualmente abbondanti.
Il numero effettivo di specie può essere derivato dalle seguenti formule:
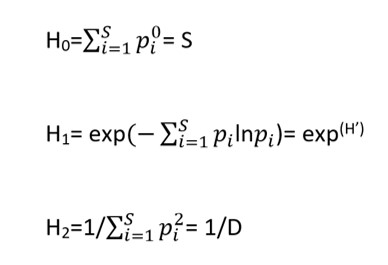
dove S è il numero di specie nel campione, pi = n/N è l’abbondanza relativa, ovvero l’abbondanza di ciascuna specie (n) nel campione divisa per l’abbondanza totale di tutte le specie nel campione (N), H’ è l’indice di diversità di Shannon e D è quello di Simpson.
La necessità di un indice universale di biodiversità assoluta ed effettiva
Nonostante questa proliferazione di indici, i precedenti tentativi di identificare una misura di diversità universale non sono riusciti a trovarne una unica per rappresentare la vera diversità di una comunità e, invece, hanno evidenziato la necessità di calcolare, in ogni studio, i valori calcolati da almeno due indici per aumentare l’affidabilità della misurazione. Pertanto, la scelta di uno degli indici effettivo di specie (H0, H1 o H2) dipende da quali aspetti della diversità dei siti di studio la ricerca intende indagare. Se si prevede che gli effetti saranno più evidenti in specie o tratti rari, allora H0 (la misura effettiva della semplice ricchezza in specie) sarebbe appropriato. Tuttavia, se si prevede che specie o i tratti dominanti saranno più importanti, allora H2 (la misura effettiva dell’indice di Simpson) sarebbe più rilevante. H1 (la misura effettiva dell’indice di Shannon) dovrebbe essere usato, invece, in situazioni in cui si prevede che specie o tratti rari e abbondanti siano ugualmente importanti.
Sebbene convertire gli indici di diversità di Shannon e Simpson in vere diversità (cioè numeri effettivi di specie) dia loro un insieme di comportamenti e proprietà comuni, la scelta di quella più appropriata potrebbe influenzare lo studio e fuorviare l’interpretazione dei risultati comparativi. Inoltre, dopo la conversione, la diversità viene sempre misurata in unità di numero di specie, indipendentemente da quale indice sia stato utilizzato, ma rappresenta solo la diversità osservata e non quella attesa (o assoluta) della comunità studiata.
Per confrontare e interpretare il numero effettivo di specie con una sola metrica e analizzare i dati sulla diversità con tecniche che non dipendono da un indice non lineare tradizionale, insieme al dott. Alfonso Monaco e al dott. Nicola Amoroso del Dipartimento di Fisica dell’Università degli Studi di Bari, abbiamo recentemente sviluppato una misura assoluta di diversità basata sul numero effettivo di specie e derivato dai numeri di Hill (H) dell’ordine 0, 1 e 2 (Cazzolla Gatti R., Amoroso N., & Monaco A. 2020. Estimating and comparing biodiversity with a single universal metric. Ecological Modelling, 424, 109020). Questo indice costituirebbe la prima misura universale, che combina sia la biodiversità assoluta sia quella effettiva correlata agli indici tradizionali più utilizzati (come S, Shannon e Simpson), in un valore numerico unico, semplice e completo che rappresenta, dunque, la diversità effettiva e assoluta (definita, AED) di qualsiasi comunità biologica.
Il nuovo indice della diversità effettiva e assoluta (AED)
Sottoforma di una diversità assoluta simile all’indice Chao (cioè specie osservate + singletons/doubletons), il nostro indice è composto dal numero effettivo di specie osservate (H0) più una proporzione tra l’esponenziale dell’indice di Shannon (H1) e l’inverso dell’indice Simpson (H2).
Il nuovo indice di diversità assoluta, costruito sul numero effettivo di specie di una comunità (AED), viene calcolato con la seguente semplice equazione:
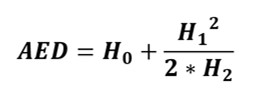
Questo nuovo indice è racchiude le informazioni dei tre principali numeri di Hill e descrive bene il profilo di diversità di una comunità, in una misura di diversità assoluta (cioè un indice simile a quello di Chao), che si è dimostrato affidabile e meno influenzato da errori di stima rispetto alle tre tradizionali misure di diversità, su cui si basa, prese da sole.
La varianza dell’AED (sottoforma di errore standard, S.E.) può essere calcolata come:
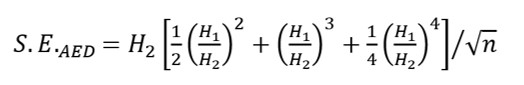
La nostra misura racchiude la ricchezza osservata e la diversità “mancante” stimata, dovuta alle specie rare e più difficili da rilevare, in un unico indice.
L’importanza delle specie rare per la biodiversità è uno dei principali interessi della conservazione biologica. La relazione tra singleton (H1 nell’indice AED) e doubletons (H2 nell’indice AED) sembra essere una misura affidabile delle “specie mancanti” in un campione. Jost (2006) ha affermato chiaramente che i tre indici di diversità di ordine zero (ricchezza di specie), uno (esponenziale dell’indice di Shannon-Wiener) e due (inverso dell’indice di Simpson) forniscono più informazioni sui campioni di qualsiasi singola misura ed è consigliabile fornire tutti e tre in ogni studio di una singola comunità. Per tale ragione è nata l’esigenza di formulare l’indice AED in grado di includere tutte e tre le principali misure di diversità in un modo che consenta non solo un calcolo della diversità effettiva di una comunità, ma anche di quella assoluta, con una metrica universale.
Un test empirico in un ecosistema marino
Dopo aver effettuato un primo test del nuovo indice su dati empirici di biodiversità forestale raccolti dal Forest Inventory and Analysis (FIA) Program degli Stati Uniti in ecosistemi temperati, boreali e tropicali, per valutarne l’affidabilità nel confrontare la ricchezza stimata di siti che hanno una grande differenza nella loro composizione specifica, abbiamo stimato la diversità assoluta ed effettiva di una comunità di barriera corallina presso la Stazione Navale Guantánamo Bay nella parte sud-orientale di Cuba da un set di dati pubblicato da Chiappone et al. (2001). In questo studio, furono raccolti dati sull’abbondanza in termini di copertura dell’area (%) per ciascuna specie di ottocoralli, coralli calcarei, alghe di fondo erboso, alghe coralline, ecc.. Abbiamo, quindi, testato le prestazioni dell’AED rispetto a quelle di altre comuni metriche di ricchezza assoluta (ovvero Chao1, Jackknife1 e 2, Bootstrapping e Michaelis-Menten) su questa comunità del reef. Come nel test sulla biodiversità forestale, anche in questo caso l’AED Index ha richiesto un basso sforzo di campionamento (una copertura dei circa il 40%) per stimare la diversità effettiva e assoluta della comunità. Inoltre, l’indice AED ha mostra una maggiore accuratezza rispetto agli altri indici nella stima, con una minore variabilità attorno alla media e nessuna sopravvalutazione e sottostima.
L’AED valuta adeguatamente la struttura della comunità ed evita le distorsioni di misurazione legate alle diverse dimensioni del campione. Confrontando l’indice AED con lo stimatore Chao1 da cui prende forma si rilevano prestazioni simili con almeno tre vantaggi principali che fanno pendere l’ago della bilancia per l’utilizzo dell’AED: rispetto all’indice Chao, la sua precisione è molto più elevata (una variabilità molto bassa attorno alla media), non mostra sovrastime e sottostime estreme ed è in grado di stimare la diversità effettiva e assoluta di una comunità calcolando solo i tre principali indici di diversità (S, Shannon e Simpson) senza la necessità di calcolare i singletons e i doubletons. In effetti, i valori degli indici di ricchezza (S), di Shannon e Simpson sono quasi sempre disponibili in letteratura (e possono essere utilizzate per produrre stime da studi pubblicati in precedenza) e sono sempre calcolabili da set di dati basati sull’abbondanza, mentre il numero di singletons e doubletons potrebbe non essere menzionato nei dataset pubblicati o essere zero nei campioni (è comune, infatti, il caso in cui singletons, doubletons o entrambi siano = 0), rendendo impossibile la stima della ricchezza assoluta delle specie dall’indice di Chao. Inoltre, gli indici del numero effettivo di specie (H0, H1 e H2) funzionano bene solo quando l’uniformità di una comunità campionata è vicina a 1 (cioè tutte le specie sono quasi ugualmente abbondanti o ugualmente rare). Tuttavia, quando l’uniformità è inferiore a 1 ed è tra 0 e 0,6, come nel caso di molte comunità naturali reali, il nuovo indice AED è maggiormente in grado di tener conto delle specie “mancanti effettive” che contribuiscono alla diversità della comunità. Infine, l’AED rappresenta uno stimatore affidabile della vera diversità di una comunità (poiché si basa sui numeri di Hill, che tengono conto anche delle abbondanze) e non solo uno stimatore della ricchezza delle specie.
Il principale punto di forza di questo nuovo indice è la sua semplicità di utilizzo per confrontare la vera diversità di comunità, ecosistemi e biomi in tutto il mondo mediante valori adimensionali (ovvero senza unità di misura), effettivi e assoluti.
C’è un mare di biodiversità ancora da misurare e proteggere, ma ora abbiamo a disposizione – finalmente – un indicatore universale che può aiutarci a farlo.
Roberto Cazzolla Gatti, Ph.D.
Biologo ambientale ed evolutivo
Professore associato presso l’Istituto di Biologia della Tomsk State University, Russia
Research Fellow presso il Konrad Lorenz Institute for Evolution and Cognition Research, Austria
Principali riferimenti bibliografici
Cazzolla Gatti, R. (2018). Biodiversity in time and space. Nova Science Publishers, New York.
Cazzolla Gatti, R., Notarnicola, C. (2018). A novel Multilevel Biodiversity Index (MBI) for combined field and satellite imagery surveys. Global Ecology and Conservation, 13, e00361.
Cazzolla Gatti R., Amoroso N., & Monaco A. 2020. Estimating and comparing biodiversity with a single universal metric. Ecological Modelling, 424, 109020
Chao, A. (1984). Non-parametric estimation of the number of classes in a population. Scand. J. Stat. 11, 265-270.
Gotelli, N. J., & Colwell, R. K. (2011). Estimating species richness. In: Biological diversity: frontiers in measurement and assessment (Magurran, A.E., McGill, B.J., Eds.), 12, 39-54.
Hill, M.O. (1973). Diversity and evenness: a unifying notation and its consequences. Ecology 54, 427–473
Jost, L. (2006). Entropy and diversity. Oikos, 113(2), 363-375.
Spellerberg, I. F., & Fedor, P. J. (2003). A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’Index. Global ecology and biogeography, 12(3), 177-179.
Simpson, E. H. (1949). Measurement of diversity. Nature, 163(4148), 688.
*Pubblicato sul numero 90 di giugno 2020 di Villaggio Globale






