Published on Theoretical Biology Forum 104 (1), 35, 2011
Author(s): Roberto Cazzolla Gatti [1]
[1] Department of Forest Resources and Environment (DISAFRI), Via San Camillo De Lellis, University of Tuscia, 01100 Viterbo, Italy
Summary: A. McFayden[1] and G.E. Hutchinson’s[2] defined a niche as a multidimensional space or ipervolume within the environment that allow an individual or a species to survive, we consider niches as a fundamental ecological variable that regulate species’ composition and relation into ecosystems. Successively the niche concept has been associated to the genetic term “fenotipe” by MacArthur[3][4] stressing the importance on what a species or a genome can show outside, either in the environmental functions or in body characteristics. Several indexes have been developed to evaluate the grade of overlapping and similarities of species’ niches, even utilizing the theory of information. However, which are the factors that determine the number of species that can coexist in a determinate environment and why a generalist species do not compete until the exclusion of the remaining species to maximize its fitness, is still quite unknown. Moreover, there are few studies and theories that clearly explain why the number of niches is so variable through ecosystems and how can several species live in the same basal niche[5][6], intended in a comprehensive sense as the range of basic conditions (temperature, humidity, food-guild, etc.).
Here I show that the number of niches in an ecosystem depends on the number of species present in a particular moment and that the species themselves allow the enhancement of niches in terms of space and number. I found that using a three-dimensional model as hypervolume and testing the theory on a Mediterranean, temperate and tropical forest ecosystem it is possible to demonstrate that each species plays a fundamental role in facilitating the colonization by other species by simply modifying the environment and exponentially increasing the available niches’ space and number. I resumed these hypothesis, after some preliminary empiric tests, in the Biodiversity-related Niches Differentiation Theory (BNDT), stressing with these definition that the process of niches differentiation is strictly addressed by species. This approach has various consequences, first in consideration of relations among species and second in terms of a better understanding of cooperation/competition dynamics.
Kauffman in the 1993 published a book titled “Self-Organization and Selection in Evolution”[7]. In this masterpiece of biology he argued that the complexity of biological systems and organisms might result as much from self-organization and far-from-equilibrium dynamics as from Darwinian natural selection. Some fellow biologists and physicists working in Kauffman’s area reserve judgment on Kauffman’s claims about self-organization and evolution. Kauffman explored complex systems ans science that focuses on the emergent and adaptive behaviors between multiple systems and their interactions, as weel as environmental, social, economic, technological systems.
Recently, Levine M.J. and HilleRisLambers[8] suggested that niches differences stabilize competitor dynamics by giving species higher per capita population growth rates when rare than where common and that coexistence occurs when these stabilizing effects of niche differences overcome species in overall competitive ability. Bastolla U. et al.[9] argued that nestedness reduces interspecific competition and enhances the number of coexisting species. These new ecological approaches conduct towards other assumptions on the role of species and their niches.
Basically, the Competitive Exclusion Principle[10] affirms that it is impossible that two species that are in competition for the same resources or ecological rules can live in the same ecosystem. This principle looks at niches as static[11] and predetermined “ecological volumes”[12] that species can fill. Mathematical models[13][14][15] and empirical analyses[16][17][18] confirm that without a clear differentiation of niches the common species become more common and rare species rarer, suggesting that niches’ differentiation strongly stabilizes the coexistence of species[19]. However, this is not a univocal relation. Thinking in terms of non-linear/non-equilibrium systems with feedback and dissipative[20] mechanisms, we can say that not only do niches enhance species coexistence but really species increase the number of niches available, thus they allow a biodiversity increment in resources exploitation within an ecosystem.
Said that, we can formulate the BNDT as follows: in natural conditions of immigration and emigration, with every environmental condition, species tend – directly or indirectly, thanks to their simple presence and life roles – to increase the number of potentially available niches for the colonization of other species, enhancing the limit imposed by the basal ipervolume, until the reach of the carrying capacity of the ecosystem. At same time, niches and mutualistic networks of the ecosystem allow, through circular and feedback mechanisms, the rise of the number of species, generating a non-linear autopoietic system[21][22].
In other words, we can argue that the species themselves, creating favourable conditions for the colonization of other species, allow the concurrent presence. This is nothing more than coexistence and the fundamental mechanism that supports the coexistence of species is the creation of diversity-related niches. The higher the number of species is, the more likely the possibilities that other species can colonize that environment are.
Trying to model the previous definition we can determine that the number of niches in a discrete time depends on the number of niches related to the initial number of species:
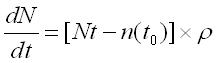
assuming that N is the number of niches, t is the time, n is the initial (at time t0) number of niches and ρ is the coefficient of niches facilitation (intended as 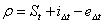 where S is the number of species at time t, i is the rate of immigration/speciation and e the rate of emigration/extinction).
where S is the number of species at time t, i is the rate of immigration/speciation and e the rate of emigration/extinction).
Solving this differential equation the number of niches at time t becomes an exponential function of the initial number of themselves elevated for the number of species multiplied for a variable c:
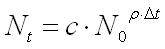
If we consider the carrying capacity (K) of the ecosystem (e), the equation can be rewritten as follows:
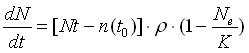
Taking discrete intervals of time, the systems (ecosystems) tend to be subjected to an increase of the number of species proportional to the number of species already present in the environment at time t, with available niches that rise in an exponential way. Systems with different initial number of species, even if they have the same chemical-physics basic conditions, after a discrete interval of time dt, will count a different number of species, directly proportional to the number of potential niches developed and to the initial number of species.
Running, instead, the model towards an infinite time (t→∞) every ecosystem, in identical chemical-physical conditions, tends to reach the same number of species that is maximum at climax and at carrying capacity level. However, as every model, it is only a simplification of reality and cannot completely represent the whole ecological roles and relations among species and the complete dynamics of niches formation. Anyway, taken with many precautions it is a easy exemplification of the theory.
Going deeper into niches formation and evolution and remembering Hutchinson[23] concept of multidimensional space, I would suggest a three-dimensional model of diversity-related niches differentiation, as showed in Fig. 1. 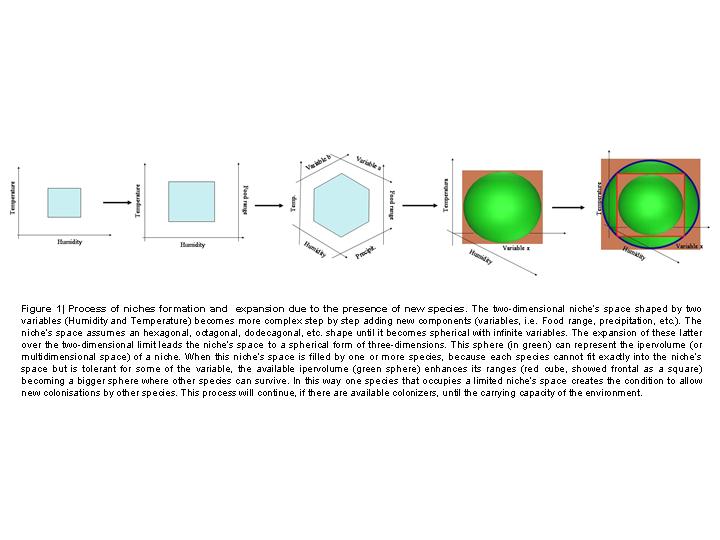 The two-dimensional system of basic variables evolves towards a three-dimensional niche’s space (a sphere) of multiple variables. When one or more species are able to fill this niche’s space, and because most of species are strict for some ecological condition but tolerant for other variables, the hypervolume (considered as the sum of every single range of variables) enhance its dimensions allowing other species to colonize the environment. In this way a niche that was originally forbidden to some species for some ecological characteristics becomes available because of the simple presence of a species that can tolerate initial conditions.
The two-dimensional system of basic variables evolves towards a three-dimensional niche’s space (a sphere) of multiple variables. When one or more species are able to fill this niche’s space, and because most of species are strict for some ecological condition but tolerant for other variables, the hypervolume (considered as the sum of every single range of variables) enhance its dimensions allowing other species to colonize the environment. In this way a niche that was originally forbidden to some species for some ecological characteristics becomes available because of the simple presence of a species that can tolerate initial conditions.
Graphically it could be imagined as a sphere that represents the basal niche’s space and a big tree (Figure 2) that fills the whole space. Roots of this tree are pioneer species (r-selected[24]), that partially are substituted during growth and trunk is one or few dominant generalist species (we can call, G-species). 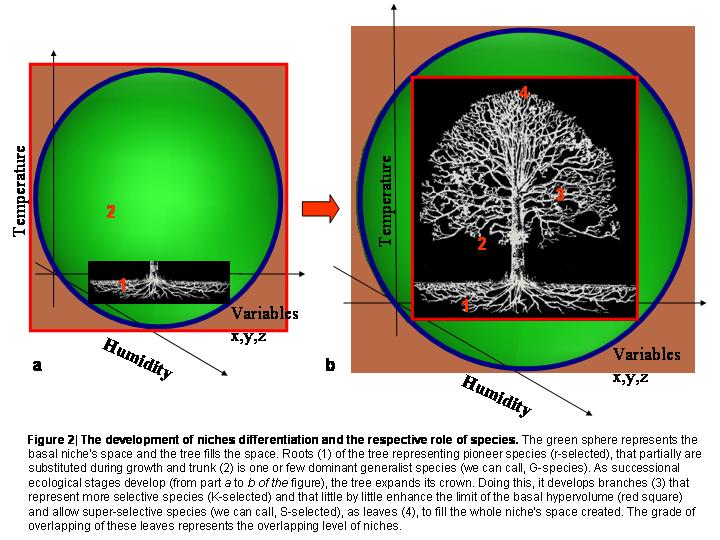 As successional ecological stages develop, the tree expands its crown. Doing this, it develops branches that represent more selective species (K-selected23) and that little by little enhance the limit of the basal hypervolume and allow super-selective species (we can call, S-selected), as leaves, to fill the whole niche’s space created. The grade of overlapping of these leaves represents the overlapping level of niches.
As successional ecological stages develop, the tree expands its crown. Doing this, it develops branches that represent more selective species (K-selected23) and that little by little enhance the limit of the basal hypervolume and allow super-selective species (we can call, S-selected), as leaves, to fill the whole niche’s space created. The grade of overlapping of these leaves represents the overlapping level of niches.
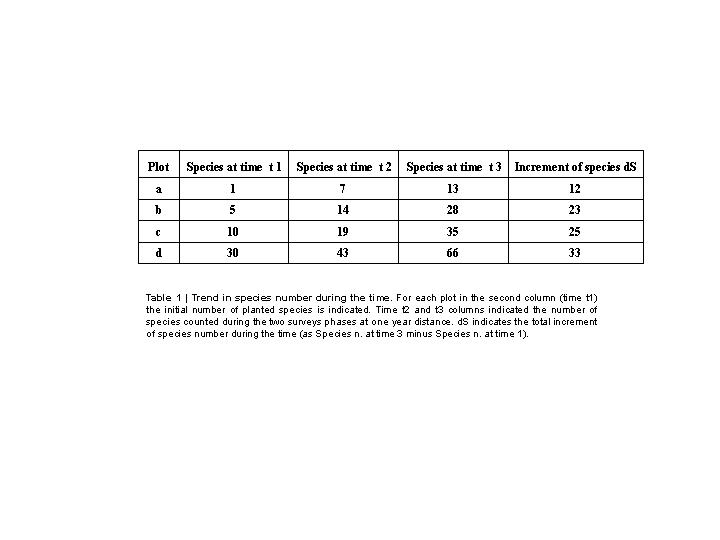
To test empirically the BNDT I realized three preliminary field analyses. The first was conducted into a Mediterranean forest ecosystem[25]. A square plot into a burned forest in the South-Italy region was selected. Four squared areas equally surrounded by wood (at same mean distance) were extracted within this plot. The number of survived species after fire (3 months before the experiment) was zero for each area. I planted respectively 1, 5, 10 and 30 endemic shrub species randomly selected, belonging to the close forest. The rate of immigration and emigration for each area was statistically the same. No speciation and extinction processes were documented due to the strict time of the survey. I came back two times, at one year distance, to count the number of species present in each sample area. The first year I found that the plot accounted respectively 7, 14, 19 and 43 species of plants and animals. The second year the number of species was respectively 13, 28, 35 and 66 (Table 1).
These results mean that the increment of species from the original situation, after two years, was of 12, 23, 25 and 33 for each sample plot, excluding the original planted species, and that this increase strictly depends on the number of initial species. The curves follow, as predicted by the model, the exponential trend (Figure 3).
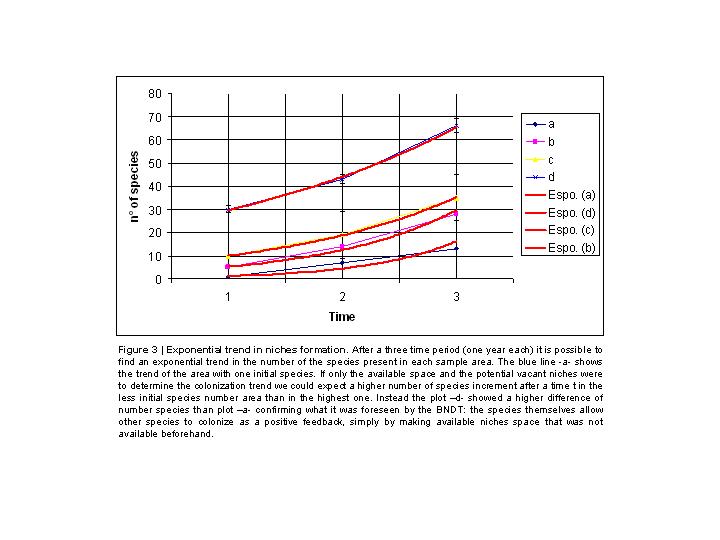 The result could appear apparently strange, particularly if we consider that in the sample area with one initial planted species there was much more potential niche’s space available to be colonized compared, for instance, to those available in the area with 30 initial species. Instead, contrarily to what expected, we found an increased number of 12 species for the first area with one initial species and 33 for the sample area with the highest number of species.
The result could appear apparently strange, particularly if we consider that in the sample area with one initial planted species there was much more potential niche’s space available to be colonized compared, for instance, to those available in the area with 30 initial species. Instead, contrarily to what expected, we found an increased number of 12 species for the first area with one initial species and 33 for the sample area with the highest number of species.
 This preliminary results show that, as predicted by the BNDT, not only the available space or the vacant niches play an important role in determining the number of colonizers but also the presence of species induces and somehow encourages the presence of others.
This preliminary results show that, as predicted by the BNDT, not only the available space or the vacant niches play an important role in determining the number of colonizers but also the presence of species induces and somehow encourages the presence of others.
Another research in Indonesia conducted last year, within the Bali Barat National Park, seems to confirm the previous results. I surveyed two different tropical forest ecosystems. One was a lowland unmanaged pristine tropical forest. The second was a selective-logged lowland tropical forest. I found that the species composition was quite similar in both forests and that the selectively logged one followed a niches development similar to the pristine one. Because in the logged forest some selected species were removed (indicated as β), we can expect a total number of species (S) differing by the close unmanaged forest (α) only in the number of logged species (S= α- β). On the contrary, the total number of species in the selectively logged forest was very low compared to what we can expected merely deducting β to α. This confirms that the removal of one species takes out many other species (>5% of the total number) from the ecosystem as predicted by the BNDT.
A third evidence of the theory can be observed in many different ecosystems like on the top of the Monte Venere, in Lazio region in Italy. In the east slope of the mountain it grows an old beech forest and in the western slope an oak forest. While oak allows a rich brushwood and the presence of vines, lianas, epiphyte and other tree species, the beech creates the condition to be quite a monocultural species. On the edge of the Monte Venere’s top It is strange to suddenly pass through a rich of biodiversity ecosystem to a monotonous sweep of one species. Considering that the climatic and environmental conditions are quite the same (even if the solar radiation and the exposure to the winds that come from Vico Lake are a bit different) it is very strange to see this situation in less than one hectare. I can suggest that the fundamental reason that lead to this phenomenon is strictly linked to the BNDT. Oak-dominated forests follow the successional evolution foreseen by the theory, where a species creates the conditions for the growth of other species rising exponentially the available niches and the limits of basal hypervolume. Beech-dominated forests instead do not seem to follow the previsions of the BNDT, mining its validity. Recently, however, has been evidenced that the Bos primigenius, before their extinction caused by human hunters, was the controlling factor of Fagus overgrowing, giving more emphasis to the role of natural history to species’ relations in shaping the ecosystems and determining the creation of new niches (trough the biodiversity-related niches differentiation mechanism) or the maintaining of a mono-species dominated environment, that frequently is the result of an ecological stress.
These fast surveys and evidences could confirm the results of the Mediterranean ecosystem plots where, because of the artificial differences in the initial number of species, the number of species at certain time periods resulted different among plots. There is necessity to better understand the validity of the BNDT in a complex tropical ecosystem and I am going to do it in the African tropical rainforest next two years.
However, theoretical and preliminary empirical approaches could lead to other questions that need to be answered: is the rule played by each species in an ecosystem indifferent in terms of niches differentiation or are generalist species more relevant to allow specialist ones to fill niches that are created in the late stages? and how does the evolution of niches change in relation to the different species that colonize the environment during the first stages?
The BNDT, following this preliminary results can be the answer to the reason why we have a higher number of species in the plot with higher initial number of species, if the immigration/emigration tax is the same in the four plots. Furthermore, a future step towards an empirical confirmation of the BNDT could be the evaluation of the behaviour of the four plots in long time to verify if the carrying capacity of the system will keep under a unique threshold the number of species living in that ecosystem, shaping the four curves in a sigmoid way to converge towards a common level of niches filled.
If these empirical evidences are consistent with the BNDT it will be possible to better understand the species’ relations and the rule they play within the ecosystems. At the same time it will be simpler to predict the behaviour of a complex system and shift our idea of niches from a static to a dynamic and biologically (biodiversity) determined vision.
More importantly, a confirmation of the BNDT validity could represent a new way to see the dominant force among species and reconsider the importance of the competitive exclusion towards a less competitive nature where one species itself, enhancing the possibilities offered by the environment, allow other species to live in the same ecosystem.
Moreover, the BNDT could offer an opportunity to link as a bridge the criticized Unified Neutral Theory of Biodiversity (UNTB)[26], that promotes the concept of irrelevancy of adaptations, with the older theory of competitive exclusion.
Finally, the BNDT increase the urgency to arrest the loss of biodiversity, because if the theory were to be confirmed right, the loss of one species could lead to the disappearance of many other species “hosted” into the niche hypervolume created by that. This dramatic evidence of the theory could take place when it is too late to revert the process.
Methods summary
To develop the BNDT I reviewed the latest theoretical, modelling and empirical approaches and extracted the common features that can adapt to real ecosystems well. The graph of the evolution of a niche space is the result of a long modelling process to better and easier represent the theory.
I followed a transect to individuate the best square plot to carry out the Mediterranean ecosystem experiment. The area is located in the region of Puglia in Italy in a semi-deciduous forest burned two years consecutively before the experiment. To be sure that no plant was present at the beginning of the experiment, not even seeds or cryptophytes, the soil beneath 10 cm deep was completely monitored and ploughed up. The 4 sample areas were of 10×10 m and formed a bigger square. To ensure that the rate of immigration/emigration was similar in each area the plots were located exactly in the middle of the forest and at the same distance from the unburned trees.
I selected after 2 years of forest survey 30 most common shrub species and I randomly selected some for each plot. To evaluate the rate of biodiversity change I counted every species present at moment of surveys in a single day for 3 times every year. Only species that can be detected by eyes (no microscopical species) were counted.
I assessed the validity of the hypothesis, confirmed by the Mediterranean plots, during a field research within the Bali Barat National Park, in Bali, Indonesia with 3 consecutive surveys that gave to the BNDT another empirical confirmation. Obviously, the short-terms of researches and the preliminary result needs to be confirmed by other studies.
References
[1] Macfadyen, A., Animal Ecology: Aims and Methods. London: Pitman (1957).
[2] Hutchinson, G.E.. A treatise on limnology v.1. Geography, Physics and Chemistry. Wiley. 1015pp. (1957)
[3] MacArthur, R. H. Population ecology of some warblers of northeastern coniferous forests. Ecology 39:599-619 (1958).
[4] MacArthur, R. H. Geographical Ecology: Patterns in the Distribution of Species (Harper & Row, 1972).
[5] Harrison, S. Local and regional diversity in a patchy landscape: native, alien, and endemic herbs on serpentine. Ecology 80, 70–80 (1999).
[6] Pachepsky, E., Taylor, T. & Jones, S. Mutualism promotes diversity and stability in a simple artificial ecosystem. Artif. Life 8, 5–24 (2002).
[7] Kauffman, S. 1993, Origins of Order: Self-Organization and Selection in Evolution, Oxford University Press, Technical monograph
[8] Levine, J. M. & HilleRisLambers, J. The importance of niches for the maintenance of species diversity. Nature 461, 254–257 (2009).
[9] Bastolla, U., Fortuna, M., Pascual-García, A., Ferrera, A., Luque, B., & Bascompte, J. The architecture of mutualistic networks minimizes competition and increases biodiversity. Nature, 458 (7241), 1018-1020 (2009).
[10] Gause, G. F. The struggle for existence. New York, NY: Hafner Publishing Company, 1934.
[11] Chesson, P. Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 31, 343–366 (2000).
[12] Chase, J. M. & Leibold, M. A. Ecological Niches: linking classical and contemporary approaches. London, UK: The University of Chicago Press, 2003.
[13] Tilman, D. Niche tradeoffs, neutrality, and community structure: a stochastic theory of resource competition, invasion, and community assembly. Proceedings of the National Academy of Sciences, USA. 101, 10854-10861 (2004).
[14] Volterra, V. Fluctuations in the abundance of a species considered mathematically. Nature 118, 558–560 (1926).
[15] Lotka, A. J. The growth of mixed populations: two species competing for a common food supply. J. Wash. Acad. Sci. 22, 461–469 (1932).
[16] Silvertown, J. Plant coexistence and the niche. Trends in Ecology and Evolution, 19, 605-611 (2004).
[17] Leibold, M. A. & McPeek, M. A. Coexistence of the niche and neutral perspectives in community ecology. Ecology 87, 1399–1410 (2006).
[18] McKane, R. B. et al. Resource-based niches provide a basis for plant species diversity and dominance in arctic tundra. Nature 415, 68–71 (2002).
[19] Adler, P. B., Hille Ris Lambers, J., & Levine, J. M. A niche for neutrality. Ecology Letters 10, 95–104 (2007).
[20] Prigogine I. and Stengers I., La Nouvelle Alliance, Gallimard, Paris (1979)
[21] Maturana, H., Everything is said by an observer. In Gaia, a Way of Knowing, edited by W. Thompson,. Lindisfarne Press, Great Barrington, MA, pp. 65-82, p. 71 (1987)..
[22] Maturana, H. and Varela, F., The Tree of Knowledge. New Science Library, Shambhala, Boston. p 242, 1988.
[23] Hutchinson, G. E. The paradox of the plankton. The American Naturalist, 95, 137–145 (1961).
[24] MacArthur, R. and Wilson, E.O. The Theory of Island Biogeography, Princeton University Press (1967).
[25] Gatti Cazzolla, R. Ambienti, flora e fauna delle Murge di sud-est, Adda Ed., Bari, 2010
[26] Hubbell, S. P. The unified neutral theory of biodiversity and biogeography. Princeton, NJ: Princeton University Press, 2001.






